|
|

|
   |
A denevérek rendje (Ordo Chiroptera) bats [english] Elterjedés és rendszertan A denevérek az egyetlen aktív repülésre képes emlőscsoport, mely képviselői evolúciójuk során rendkívül változatos testfelépítésre, morfológiára és ökológiai sajátosságokra tettek szert. Változatosságuk egyik kiugró példája a rend legnagyobb és legkissebb képviselője közötti méretbeli különbség: míg a Délkelet-Ázsiában elterjedt dongódenevér (Craseonycteris thonglongyai) csupán 3 gramm tömegű, addíg a Fülöp-szigeteki Acerodon jubatus repülőkutya 1.500 gramm és másfél méter szárnyfesztávolságú. A jelenleg leírt több mint 1.100 faj 19 családot alkot, ezáltal az emlősök (Mammalia) legfajgazdagabb rendjét képezi a rágcsálók (Rodentia) után. Az Északi- és Déli sark területein valamint a sivatagokon kívül világviszonylatban mindenhol megtalálhatóak, de ahogyan sok más állat és növénycsoport esetében, a legnagyobb fajdiverzitás a trópusokon található.
A denevérek rendjének változatosságát tükröző rajz, Ernst Haeckel 1904-es „Kunstformen der Natur” című művéből. Megfigyelhető az orrfüggelékek és a fülszerkezet nagyfokú diverzitása. Klasszikus értelemben két alrendet különböztetunk meg: Testfelépítés és speciális érzékszervek A denevérek testfelépítése a repülő életmódnak, tehát az aerodinamikai törvényeknek megfelelően fejlődött ki. A rend latin eredetű elnevezése („chiroptera”, cheir - kéz, pteron - szárny) is utal a jellegzetes testfelépítésre. Testük karcsú, kúp alakú, súlypontja a test melltájékán helyezkedik el. A bordák elcsontosodtak és szilárdan kapcsolódnak a szegycsonthoz, valamint a csigolyákhoz. Nyaktájon a gerincoszlop erősen kifelé ívelt, ezáltal lehetővé vált, hogy a mellüreg befogadja az igen terjedelmes és nagy teljesítőképességű szívet. A vállövben található a repülésben szerepet játszó izmok legnagyobb része.
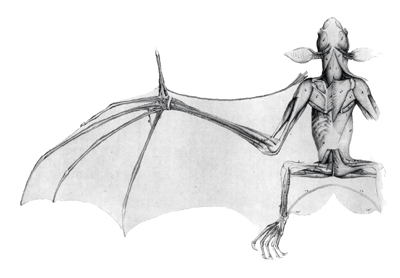
A denevérek általános felépítését jól tükröző illusztráció, George Cuvier „Leçons d'Anatomie Comparée” 1800-ban született munkájából. Megfigyelhető a „szárny” felépítése, a többszörösen megnyúlt ujjpercek, illetve a közöttük feszülő bőrvitorla. Felépítését tekintve a denevérek szárnya minden fő részében megfelel az emlősök felkar-, alkar- és kéztájék-szerkezetének, csupán arányaikban és funkciójukban térnek el. A repülő életmódnak megfelelően az elülső végtagok számos módosuláson mentek át, melyek főképp az alkar és ujjcsontok megnyúlásával hozhatók összefüggésbe. Ezen megnyúlt csontok között, valamint a testhez és lábakhoz tapadva feszül a repüléshez elengedhetetlen bőrvitorla (patagium). Elhelyezkedése alapján több részét különböztetjük meg. A felkar és alkar elülső éle mentén feszül az elővitorla vagy szélfogó (propatagium). E mögött, a kar és a láb között található a karvitorla vagy öregvitorla (plagiopatagium). Az első és ötödik kézujj között húzódik a kézvitorla (dactylopatagium). Végül, a lábak és a farok között található bőrfelület a zsákmány megszerzésében is szerepet játszó farokvitorla (uropatagium). A szárnymorfológia az illető faj vadászterületének függvényében alakult ki. Fajok, melyek sürű vegetációban vadásznak, rövid, széles szárnyakkal rendelkeznek, a jobb manőverezés érdekében (pld. nagy patkósdenevér, szürke hosszúfülű denevér). Azon fajok, melyek nyílt, vegetáció nélküli térségekben vadásznak, hosszú, keskeny szárnyakkal rendelkeznek (pld. rőt koraidenevér, óriás koraidenevér). Ez a felépítés elősegíti a gyors repulést is. A mellen és a felkar felső részén helyezkednek el azok a nagyméretű izmok, melyek erőteljes munkája által valósul meg a szárnyak mozgatása.
Akrobata módjára, egylábon függeszkedő nagy patkósdenevér hibernálás közben, mely függeszkedés azonban semmilyen nemű energia befektetéssel nem jár az egyed számára. A denevérek testét bunda, szőrzet fedi, mely kiegyenlítve testfelületük egyenetlenségeit, aerdinamikus formát kölcsönöz az állatnak, illetve hőszigetelő szerepet is betölt. Négyféle szőrtípust különböztethetunk meg. A fedőszőrök hosszú, egyenesek vagy íveltek, és erősen pigmentáltak. A szintén pigmentált koronaszőrök hosszú, erős szálú, enyhén ívelt szőrök, végükön ellaposodnak. A fedőszőrökkel együtt alkotják a felszőrzetet. Az alszőrzet a pihe és gyapjúszőrökből all. A különböző mechanikus ingerekre érzékeny szinuszszőrök és tapintószőrök az állat pofáján találhatóak. A mintázottság ritka jelleg a denevérek bundája esetében, leginkább az egyszerű szürke, barna vagy esetleg ezek kombinációi ismeretesek. A vedlés évente egyszer, tavasszal történik. Tájékozódás és táplálékszerzés ultrahangok segítségével A denevérek tájékozódása hosszú ideig ismeretlen volt a tudomány számára. Azonban Lazarro Spallanzani 1793-as és Donald Griffin 1938-as kísérletei fény derítettek arra, hogy a denevérek ún. echlokációval, azaz különböző tárgyak felületéröl visszaverődő hanghullámok segítségével tájékozódnak. Tulajdonképpen egy biológiai radarról van szó.
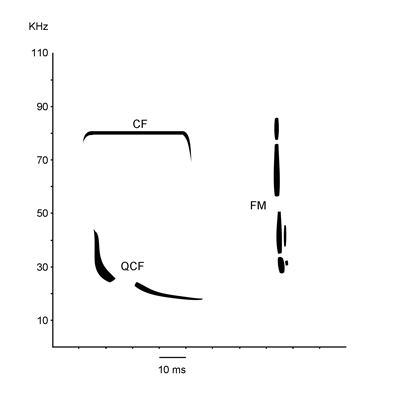
A denevérek által kibocsájtott ultrahangok három alapvető típusa. CF (constant frequency) – állandó frekvenciájú (pld. nagypatkós denevér), QCF (quasi-constant frequency) – majdnem állandó frekvenciájú (pld. korai denevérek), FM (frequency modulated) – változó frekvenciájú (pld. hosszúfülű denevérek). A három alaptípuson kívűl léteznek ezek kombinációi is, melyekben az egyik összetevő (az állandó vagy a változó frekvencia) dominál (pld. FM-qcf). A táplálékszerzés problémáját a denevérek három elkülöníthető szakaszban valósítják meg. Az első szakasz a detektálás: a denevér ellenőrzi, hogy jelen van vagy nincs a saját visszavert ultrahangjelzése, amiből aztán az esetleges zsákmányállat jelenlétére vagy hiányára következtet. A második szakasz az osztályozás szakasza, ahol a célról visszaverődő hanghullámok információt szolgáltatnak a cél nagyságáról, alakjárol és állagáról. Ez a hullámok ritmusos frekvencia- és amplitudó-váltásában nyilvánul meg. A harmadik szakasz a lokalizáció szakasza, ahol sor kerül a kiválasztott cél koordinátáinak (magasság, sebesség, távolság, irány) meghatározására, mely szakasz a tulajdonképpeni zsákmányolással ér véget. Az ultrahangos tájékozódáson kívűl a denevérek képesek vizuális és/vagy olfaktikus ingerek után is tájékozódni, főképp a trópusi fajok. A hazai fajok szemeit általában szőrzet takarja, de látásuk elegendő az alkonyati fényben való tájékozódáshoz. Szaglás alapján választják ki egyes fajok (pld. közönséges denevér) az étrendjüknek megfelelő rovarfajokat, de a rovar által kibocsájtott zaj is fontos szerepet játszik a táplálékszerzésben. A koponya formája összefüggést mutat a táplálkozás módjával, mely a fogak felépítése alapján észlelhető. A denevérek fogazata heterodont és diphydont. Különlegesen jellegzetes a zápfogak háromosztatú és daraboló szerkezete. A felső és alsó állkapocs zápfogai oly módon záródnak, hogy a tarajok egymás melletti elcsúszása a rovarok kitinpáncéljának feldarabolását segíti elő. A denevérek agyszerkezete hűen tükrözi azon átalakulásokat, melyek a repüléssel, tájékozódással és táplálkozással hozhatóak kapcsolatba. A legtöbb kisdenevér szaglólebenyében visszafejlődés következett be, ezzel szemben az agyvelő hátsó része gazdagon redőzött. Itt található a hallópálya központja, ugyanakkor itt valósul meg a bonyolult repülési manőverek összehangolt irányítása is. A hátsó ikertestek, melyek a hang utáni tájékozódásban játszanak szerepet, sokkal fejlettebbek, mint a látásban szerepet játszó elűlső ikertestek. A denevérek emésztőszerve igen rövid és csak kis mértékben differenciálódott. Ez a gyors és hatásos metabolizmus, valamint a repüléshez való alkalmazkodás következtében alakult ki. Vándorlás Akárcsak a madarakra, a denevérekre is jellemző az őszi és tavaszi vándorlás. Az őszi vándorlás az ideális telelőhely felé irányul, melyek általában barlangok és bányajáratok, vagy más, stabil hőmérsékletet és zavartalanságot biztosító helyek. A tavaszi vándorlás a szülőkolóniák irányába törtenik, ahol a nőstények világra hozhatják utódaikat. A megtett táv nagy intervallumok között mozog, pár 10 km-től több 100, vagy akár 1000 km-es is lehet. A vándorlás, illetve a megtett táv szempontjából a denevéreket három csoportba lehet osztani: Szaporodás A denevérek, más állatoktól eltérően, ősszel párzanak. Ebben a periódusban, augusztus közepétől október elejéig, éjszakánként párzóhelyeket, elsősorban barlangokat keresnek fel, ahol esetlegesen más kolóniákból származó egyedekkel kerülnek kapcsolatba. A hímek egyetlen nap erejéig 5-10 nőstényből álló háremet alakítanak ki, melyekkel párosodnak. A következő éjjel folyamán a hárem összetétele megváltozik. A párzási időszak a tél kezdetéig elhúzódhat, de a pete megtermékenyítese csak tavasszal következik be, amikor a táplálékbőség lehetőve teszi az embrió megfelelő fejlődését. Ez alól csak a hosszúszárnyú denevér kivétel, melynél a pete már ősszel megtermékenyül, de az osztódás kezdeti stádiumában megáll, és csak tavasszal folytatódik fejlődése. Ennek megfelelően a hosszúszárnyú denevér vemhességi időtartama 7 hónap, míg többi fajé csupán 1.5-3 hónap. A vemhesség időtartama két héttel is megnőhet kedvezőtlen időjárási körülmények között.
Közönséges denevérekből álló szülőkolónia a Nyugati Szigethegységben (fotó: Barti L.). A jobb oldali fotón megfigyelhető a juvenilis egyedek nagyfokú parazitáltsága. A hibernálási periódus (lásd alább) vége felé közeledve, a tavasz érkezésével gyakoribbá válnak az ébredések és a barlang előtti tájékozódó repülések. Március végén a telelő kolóniák feloszlanak, a vemhes nőstények pedig a nyári szálláshelyekre vonulnak. A nőstények ébredése után megtörténik a petesejt megtermékenyítése és megindul az embrió fejlődése. Szülés közeledtével a nőstények ún. szülőszobákba rendeződnek, szülőkolóniákat alkotva. Ebben a periódusban a hímek izoláltan élnek, általában távol a szülőkolónia helyétől. Fajtől és időjárástól függően a denevérek május végén, június elején hozzák világra utódaikat. Míg kezdetben a nőstények a bundájukba és emlőjükbe kapaszkodó kölyköket is magukkal viszik táplálékszerzési útjaikra, addig egy hét elteltével már eléggé fejletté válnak ahhoz, hogy a szálláshelyen hagyják őket. A kölykök 6-8 hét után válnak röpképessé. Ivarérettségüket a nőstények 3-4 hónap, míg a hímek másfél éves korukban érik el. A denevérek K-strategisták, általában egy, ritkább esetben két utódot hoznak a világra. A többi kisemlőssel ellentétben szokatlanul hosszú ideig élnek, életkoruk évtizedekben mérhető. A hibernálás A legtöbb denevérfaj az éjszaka folyamán keresi, illetve szerzi meg táplálékát, a nappal közeledtével pedig ún. letargikus állapotba kerül. Ez az állapot a normális, aktív testhőmérséklet csökkenését jelenti, melyet a környezet hőmérsékletének növekedése váltja ki, szerepe pedig az energiával való takarékoskodás. A testhőmérséklet csökkenése fokozatosan és szabályzott módon történik. Pld. a rőt koraidenevér esetében a testhőmérséklet 80 perc alatt 38.8 °C-ról 18 °C-ra csökken. A tél közeledtével a letargikus állapot egyre gyakrabban és egyre nagyobb intenzitással (mélységgel) fordul elő, induktorai a hőmérséklet és a táplálékmennyiség csökkenése. Maga a folyamat egy több naptól több hétig, hónapig terjedő időintervallumot ölel fel, mely alatt az állat testhőmérséklete drasztikusan lecsökkenhet, esetlegesen egészen a fagypont közeléig. A hibernálás szerepe a táplálékszegény periódusok átvészelése. Ennek előfeltétele, hogy az állat a meleg periódusokban elegendő zsírmennyiséget halmozzon fel, melynek mennyisége a hibernálás idejére elérheti a testsúly 20-30 százalékát. A felhalmozási folyamat a késő nyári hónapokban kezdődik és kedvező időjárási feltételek mellett november közepéig tart. A raktározási szakasz idejére a napi mély letargikus állapotnak tulajdoníthatóan csökken az energiafelhasználás, ezáltal a begyűjtött táplálék nagy része télire raktározódik el. Megjelölt denevérek viselkedését tanulmányozva megállapították, hogy az állatok redundáns hibernálás közben több alkalommal is felébrednek és pihenőhelyüket változtatják. A ébredési idő hossza függ a téli álom mélységétől és a külső hőmérséklettől is. Egy 4-6 g testtömegű közönséges törpedenevér átlagosan 30 perc alatt ébred fel, míg a jóval nagyobb testtömegű közönséges denevérnek 70 percre van szüksége a testhőmérséklet 5 °C-ról 38 °C-ra való emeléséhez. A felébredések gyakoriságát leginkább a hőmérséklet változása befolyásolja, de zavarás esetén is végbemehet. A kistermetű denevérfajokra jellemző, hogy kevesebb alkalommal ébrednek fel és alacsonyabb hőmérsékleten hibernálnak mint nagyobb termetű rokonfajaik. A tél kezdetével egyre ritkábban kerül sor periodikus ébredésre, de a tavasz közeledtével újból gyakorivá válnak. Az időjárási viszonyok kedvező alakulása esetén (a tavasz közeledtével) a denevérek elhagyják téli szálláshelyeiket.
Párakicsapódás hibernáló denevéren. A barlangi környezet magas páratartalma lehetővé teszi, hogy a denevérek a téli periódus folyamán kevesebb vizet veszítsenek, megakadályozva ezáltal a kiszáradást. A denevérek telelőhelyeiket a következő feltételekhez kötötten választjak meg: a barlang zavarástól mentes legyen, megfelelő mennyiségű kapaszkodásra alkalmas felület legyen jelen, 2-10 °C közötti átlaghőmérséklet uralkodjon, illeve szükséges egy magas páratartalom (70-100%) és barlangbeli légáramlatok jelenléte. A zavarásmentes közegek a kevesebb alkalommal történő felébredést teszik lehetővé. Általános értelemben minél zavarásmentesebb egy barlangi élőhely, annál ideálisabb hibernálohelyet biztosít a denevérek számára. A különböző fajok más és más típusú zavarásokra reagálnak. Egyes fajok tapintási ingerekre is felébrednek, míg mások fény-, hang-, és hőmérsékleti ingadozások okozta ingerekre reagálnak. A gyakran használt telelőhelyek átlagos hőmérséklete a 2-10 °C-os tartományba esik. A különböző fajok különböző hőmérsékleti preferenciákkal rendelkeznek (pld. nyugati piszedenevér 0-4°C, nagy patkósdenevér 7-10°C), de ez nem zárja ki annak a lehetőségét, hogy az illető faj más hőmérsékleten is hibernáljon. Vannak fajok melyek kevésbe függenek a hőmérséklettől és nagyobb mértékű változásokat is elviselnek. Pld. a kis patkósdenevér ugyanabban a barlangban megtalálható a bejárati hidegebb és változékonyabb szakaszokban, mint a barlang belső, magasabb és állandó hőmérsékletű szakaszaiban.
Hibernáló hosszúszárnyú denevérek a Királyerdő hegységben. Az állatok csoportos elhelyezkedése elősegíti egy stabil hőmérsékelt fenntartását, illetve jelentős mértékben csökkenti a szervezet vízveszteségét. Főképp a barlanglakó fajok esetében, melyek gyorsan veszítik szervezetükbol a vizet, fontos hogy a kiválasztott telelőhely magas páratartalommal rendelkezzen melynek érteke 70-100% között kell legyen. Ellenkező esetben a hibernáló állatok metabolizmusa nem tud megfelelni a körülményeknek, az állatok deszhidratálódnak és elpusztulnak. A barlangra jellemző esetleges légáramlatok, abban az esetben, ha a barlang két nyílással rendelkezik (bejárat-kijárat), káros hatással lehetnek a denevérekre. Ebben az esetben a külső környezet hőmérséklet-változásai jobban éreztetik hatásukat, és nagyobb a hőmérsékletingadozás. Ennek elkerülése végett a denevérek leggyakrabban olyan barlangokat választanak telelőhelyül, amelyeknek egyetlen nyílása van, vagy zsák alakúak. Románia denevérfaunájában jelen vannak olyan fajok melyek izoláltan, magányosan hibernálnak (pld. nyugati piszedenevér, barna hosszúfülű denevér), illetve léteznek olyan fajok melyek csoportosan telelnek (pld. hosszúszárnyú denevér). Ezen csoportosulások nagysága a pár 10 egyedtől több 1.000 (esetleg több 10.000) egyedig terjedhet. Végul léteznek olyan fajok, melyek hibernálhatnak úgy izoláltan, mint csoportosan. A nagy patkósdenevér esetében az egyedek csoportokba tömörülnek, de az egyedek testfelülete nem érintkezik. Minden esetben a tömörülés oka a felmelegedés, mivel csoportosulásuk által háromszorosan csökkenthetik a testük és a környező levegő közötti kontaktust. A Romániában honos denevéreket pihenő- illetve hibernálóhely-preferenciájuk alapján három alapvető csoportba lehet elkülöníteni. Megkülönböztetünk fitofil, litofil és antropofil fajokat. A fitofil (odúlakó) fajok fák odvaiban, fakéreg alatt, fák hasadékaiban pihennek (pld. barna hosszúfülű denevér, durvavitorlájú denevér, közönséges törpedenevér, rőt koraidenevér, nyugati piszedenevér). A litofil (barlangi) fajok barlangokban, sziklahasadékokban, elhagyott bányákban, pincékben, hidak bolthajlásaiban pihennek (pld. kereknyergű patkósdenevér, hosszúszárnyú denevér, nagypatkós denevér, közönséges denevér). Az antropofil fajok emberi településeken található búvóhelyeket választanak szálláshelyül: padlásokat, ereszhasadékokat, panelek illesztéseit (pld. szürke hosszúfülű denevér, közönséges késeidenevér, csonkafülű denevér). Ezt az osztályozást azonban nem lehet teljesnek tekinteni, mivel az egyes fajok tavaszi, nyári szálláshelyeiktől függetlenul a téli periódusban barlangokban találják meg az ideális hibernálási körülményeket. A denevérek evolúciója A földtörténet mintegy 70 millió évvel ezelőtti, Kréta és Paleocén közötti periódusában számos példát találunk az élővilág diverzitásának növekedésére. Ekkor kerül sor a dinoszauruszok eltünésére, illetve a növények, rovarok és emlősök robbanásszerű feljődésére is. A denevérek fejlődéstörténetének első képviselője az Észak Amerikai Icaronycteris index, mely megközelítőleg 51 millió éves és meglepő módon már kifejlett denevér jellegekkel rendelkezik (aktív repülés, ultrahangos tájékozódás). További fosszilis maradványokat találtak Európa, Afrika és Ausztrália lelőhelyein (pld. Paleochiropteryx, Archeonycteris, stb.). Azonban a denevérek evolúciójának két kulcslépését (a repülés és az ultrahangos tájékozódás megjelenése), köztes alakok hiányában csak elméleti szinten lehet meghatározni. A többi egyedi denevérjelleg (koloniális és éjszakai életmód, hibernálás) valószínűleg a repülés kialakulása után ment végbe. A létező (számos) elmélet alapján több forgatókönyv lehetséges, melyek főképp a két fő denevértulajdonság (repülés és ultrahangos tájékozádás) megjelenési sorrendjét próbálják tisztázni. Egyes elméletek szerint először az aktív repülés jelent meg (fákról, ágakról való egyre nagyobb ugrásokból kifejlődve), míg az ultrahangos tájékozódás/táplálékszerzés csupán követte ezen jelleget, kihasználva az új környezet (légtér, repülés) által nyújtott lehetőségeket. Más elméletek alapján az proto-denevér faágakról függeszkedve, passzív módon, de ultrahangok segítségével kereste táplálékát, majd lokalizálva azt, egyre nagyobb ugrásokkal (később már repülve) próbálta megszerezni azt. De léteznek elméletek, melyek a két tulajdonság párhuzamos evolúciójának hívei. A többi denevér jelleg (koloniális és éjszakai életmód, hibernálás, széles táplálékspektrum) kialakulása tovább bonyolítja az evolúciós képes. Azonban ez a kép sosem lesz tiszta, míg nem kerülnek a felszínre fosszilis denevérek, melyek a különböző evolóciós megvalósítások köztes lépéseit/jellegeit mutatják. Ajánlott romániai irodalom - Decu V., Murariu D., Gheorghiu V. (2003): Chiropterele din România. Ghid instructiv şi educativ. Institutul de Speologie „Emil Racoviţă” al Academiei Române – Muzeul Naţional de Istorie naturală „Grigore Antipa”, Editura Art Group SRL, Bucureşti, pp. 521. - Borda D., Borda C. (2005): Chiropterele. Relaţii cu omul şi mediul înconjurător (Bats – Relations with environment and us. Editura Napoca Star, Cluj Napoca, pp. 164. - Done T., Valenciuc N. (2001): Liliecii: între Mit şi Adevăr. Editura Fundaţia de Speologie "Club Speologic Bucovina", Suceava, pp. 108. - Valenciuc N. (2002): Chiroptera, Mammalia, Fauna României, vol. XVI., fasc. 3. Editura Academiei Române, Bucureşti, pp. 171. Ajánlott nemzetközi irodalom - Altringham J. D. (1998): Bats: Biology and Behaviour. Oxford University Press, Oxford. - Bihari Z. (1996): Denevérhatározó és denevérvédelem. Magyar Madártani és Természetvédelmi Egyesület, Budapest. - Dietz C., von Helversen O., Nill D. (2007): Die Fledermäuse Europas und Nordwestafrikas. Biologie, Kennzeichen, Gefährdung. Kosmos Verlag, Stuttgart. - Kunz T. H. (1982): Ecology of Bats. Plenum Press, New York - Kunz T. H., Fenton M. B. (2003): Bat Ecology. The University of Chicago Press, Chicago. - Kunz T. H., Racey P. A. (1999): Bat Biology and Conservation. Smithsonian Institution Press, Washington. - Mitchell-Jones A. J., McLeish A. P. (2004): 3rd Edition Bat Workers Manual. Joint Nature Conservation Committee. - Simmons N. B. (2005): An Eocene Big Bang for bats. Science 307: 527-528 - Simmons N. B., Seymour K. L., Habersetzer J., Gunnell G. F. (2008): Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation. Nature 451: 818-821 - Wilson D. E., Reeder D. M. (2005): Mammal Species of the World. A Taxonomic and Geographic Reference, Johns Hopkins University Press, Baltimore. |
|